真的想問:淡水魚被衝入大海後去哪了?還能活嗎?
你以爲的黃河:

來源:自己畫的
實際上的黃河:
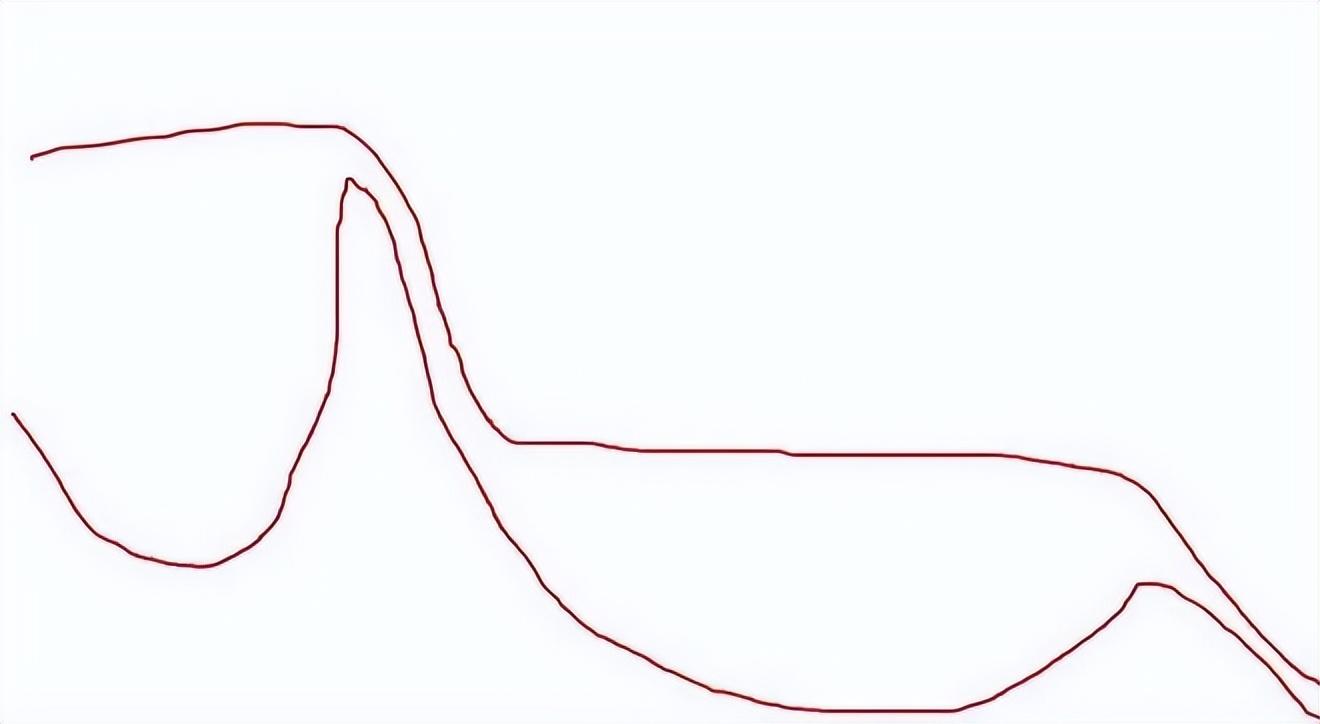
來源:自己畫的。鼠標畫得有點抽象,意思到了就行。
網上有人疑惑,黃河每天把魚衝入大海,淡水魚在海里無法存活,那麼,這些淡水魚最後去哪了呢?
之所以有這個疑問,可能是很多人以爲黃河就像“直腸子”,魚總是能被衝到大海里去。但看了上面兩張圖,相信大家應該已經瞭解,其實不是這樣的。今天,我們就來詳細聊聊這個話題。
黃河並不是直腸子
由於水流緩急、高低落差,以及地質硬軟等因素的不同,水流在不同地區帶走的泥沙體量不同,因此江河會形成諸多天然的水下深潭。
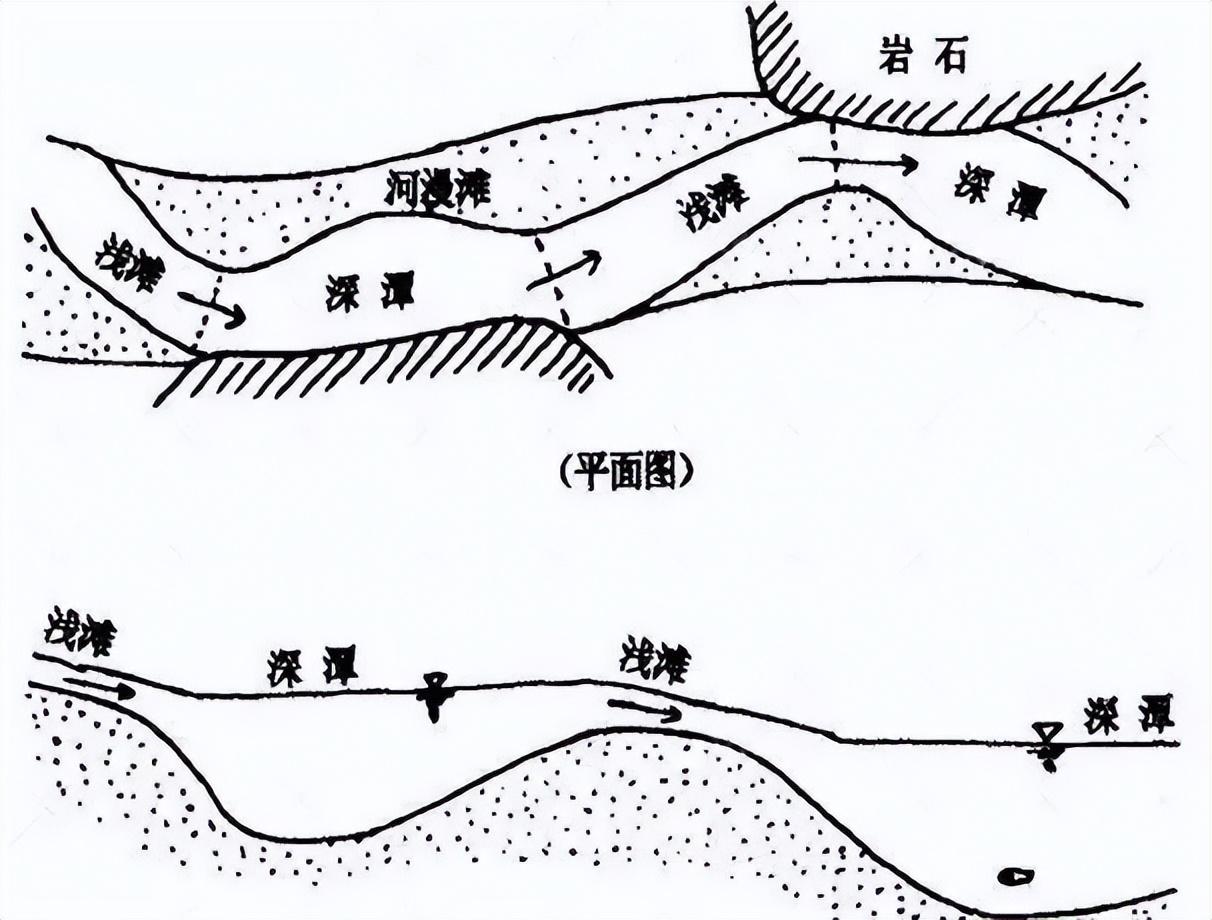
來源:河道治理 500 例

來源:網絡
在地勢開闊地帶,還能形成大湖。
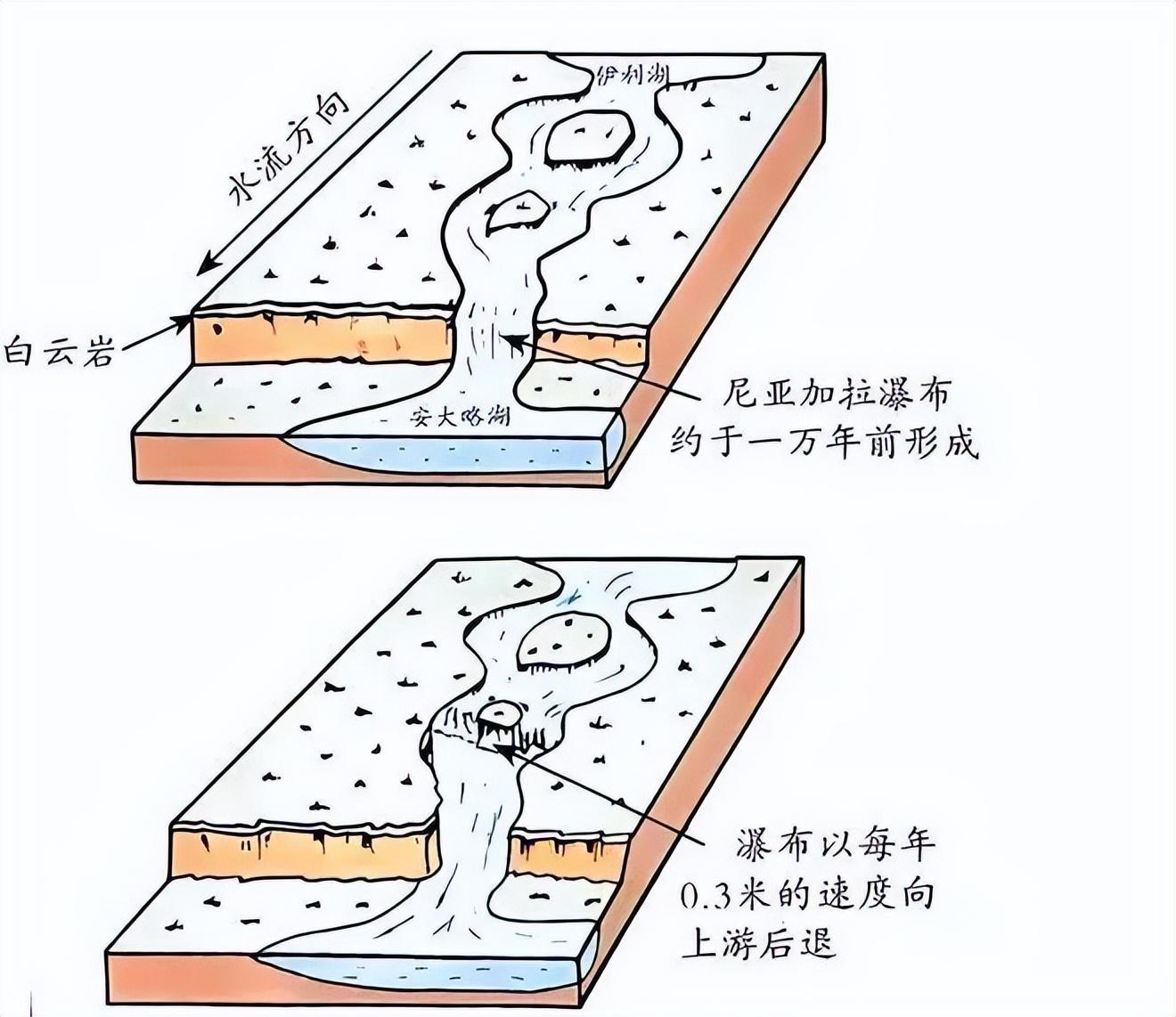
來源:網絡
作爲脊椎動物,魚類也是需要睡覺、休息的。
如果黃河真的是開篇第一張圖那樣,全程都只有持續向下的水流,的確很難會有魚可以生存。即便有,也只能是在有大量亂石和水草的地方。
有過農村小溪捕魚經驗的人就會知道,有固定魚羣的地方,基本上只有水坑和窪地。

深潭和有水草的淺灘更容易有魚。來源:河道治理 500 例
一條蜿蜒而行的江河,因爲江灘、河灘的隨機分佈,也使得水流流速有着很大的變化。
流水更緩的區域不僅往往水草更加豐茂,水陸交接生態複雜,也會有更多的固定魚羣。
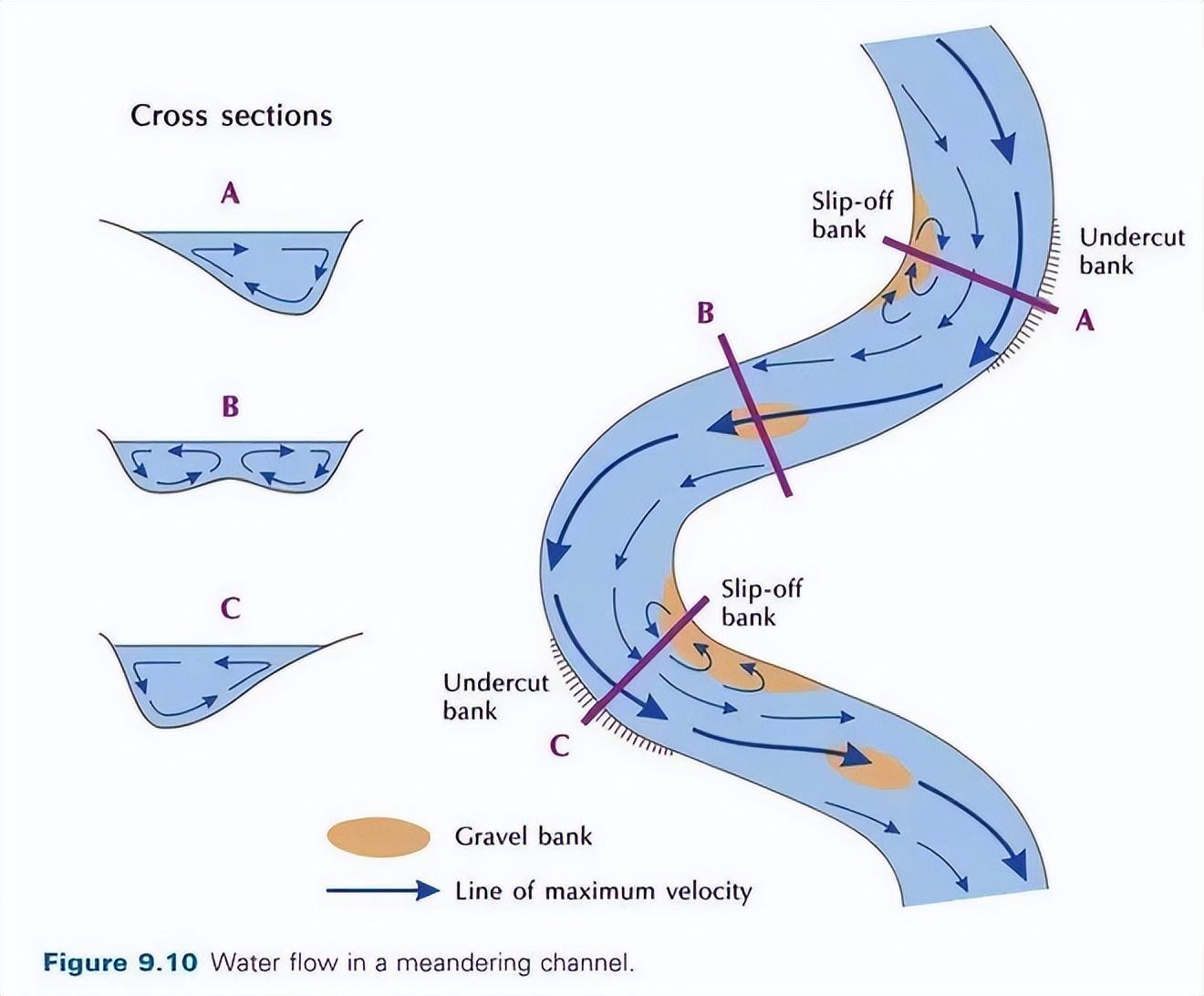
來源:upraveno podle HUGGET, 2007
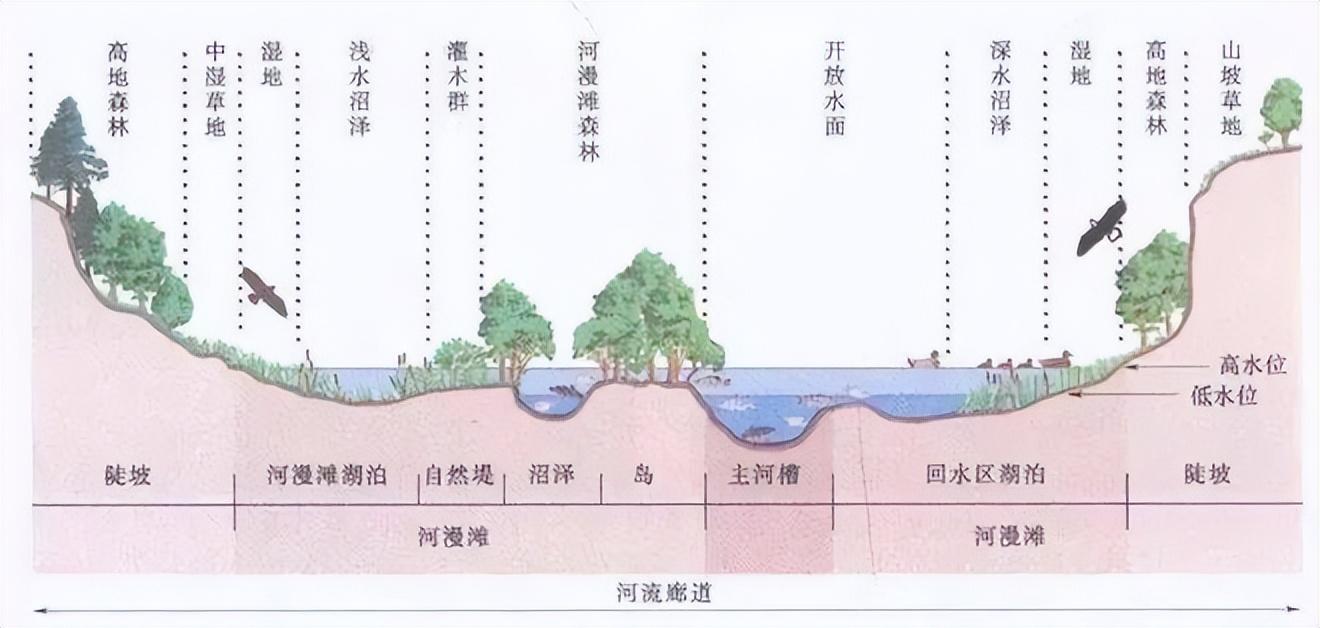
來源:FISRWG,1998,中文網絡漢化版
稍微有點急的溪流,就很難會有魚羣在固定的地方出現。
即便有魚出現,要麼很快被衝往更下游的窪地,要麼力爭上游,來到更上游的窪地。
正是因爲淡水魚的生存環境,是一個一個通過湍急水流連接的窪地,所以淡水魚才演化出了逆水而遊的習性。
每次大雨,上游都會形成更大的或者新的窪地。
一般小魚是很容易被水流沖走的,但上游水流更緩,更小,不僅小魚不容易被沖走,即便被沖走,也還會遇到很多窪地,有更多的生存機會。
基本上中小型魚類,都會在大雨、洪水中有逆水而遊的習性。
但其實,衝回來的魚居多。小時候有過捉魚經驗的都知道,大雨的時候,哪怕拿個竹兜放在小溪中,都能接到從下游池塘一路游上來,然後力竭再次被衝下來的魚。
能到上游的魚越少,也意味着到達之後,也越具有生存優勢。這裏不僅食物多,蟲子、植物果實豐富,而且往往還缺少天敵。除了基本的生存外,在上游產卵、生育後代,也會更有優勢。
總之,江河複雜的地理環境決定了,大量的魚並不會直接被衝到海里去。

蜿蜒曲折的河流。圖庫版權圖片,轉載使用可能引發版權糾紛
當然,江河下游,也生存着很多魚,最終被衝到海里的魚也不是沒有。
那麼,被衝到海里的魚會怎麼樣呢?
被衝到海里的淡水魚
後來怎麼樣了?
1
當然是游回去
我們知道江河入海口,也是徑流最大的地方。但這裏,由於水域開闊,水流速度反而比較慢。
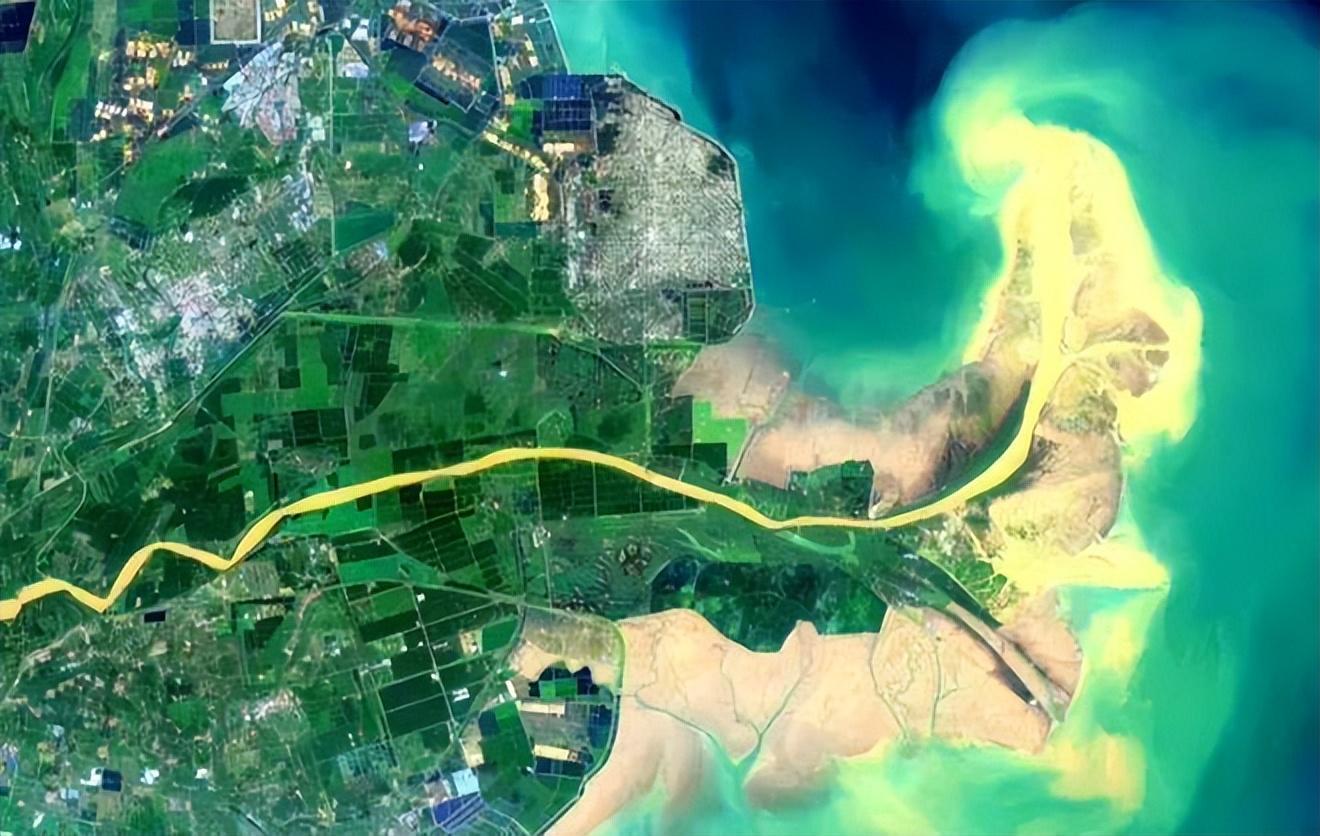
黃河入海口(來源:山東東營黃河入海口衛星圖,8.14)
絕大多數誤入入海口的魚類,都有能力再次游回去。
而江水河水與海水也不是涇渭分明的,而是存在一個特殊的鹽度緩衝區[1]。鹽度從近岸的 0.1% 漸變到外海的 3%,形成一道天然的過渡帶。
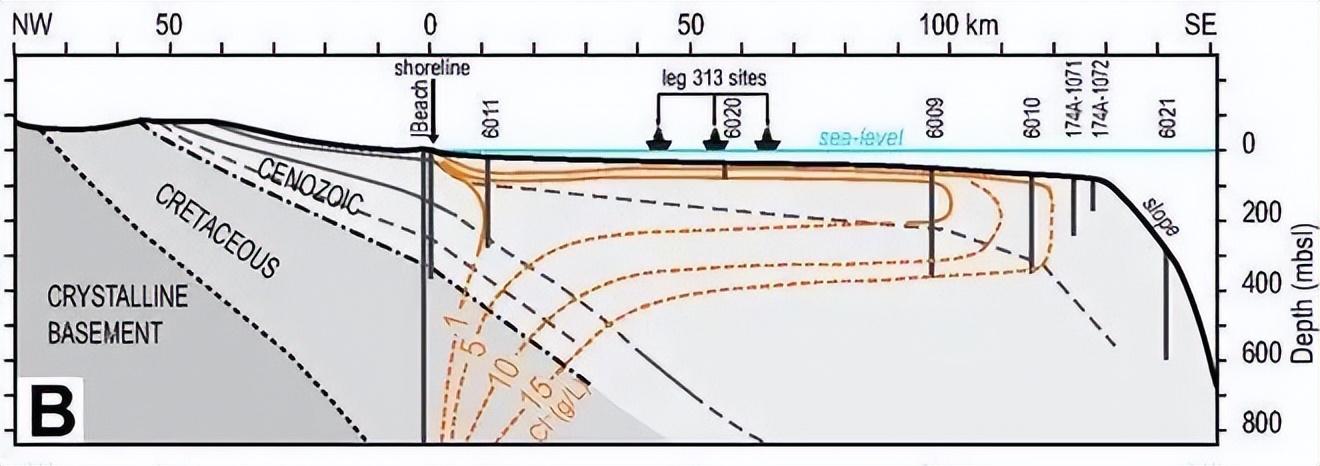
入海口的典型鹽度梯度變化(來源:文獻[1] )
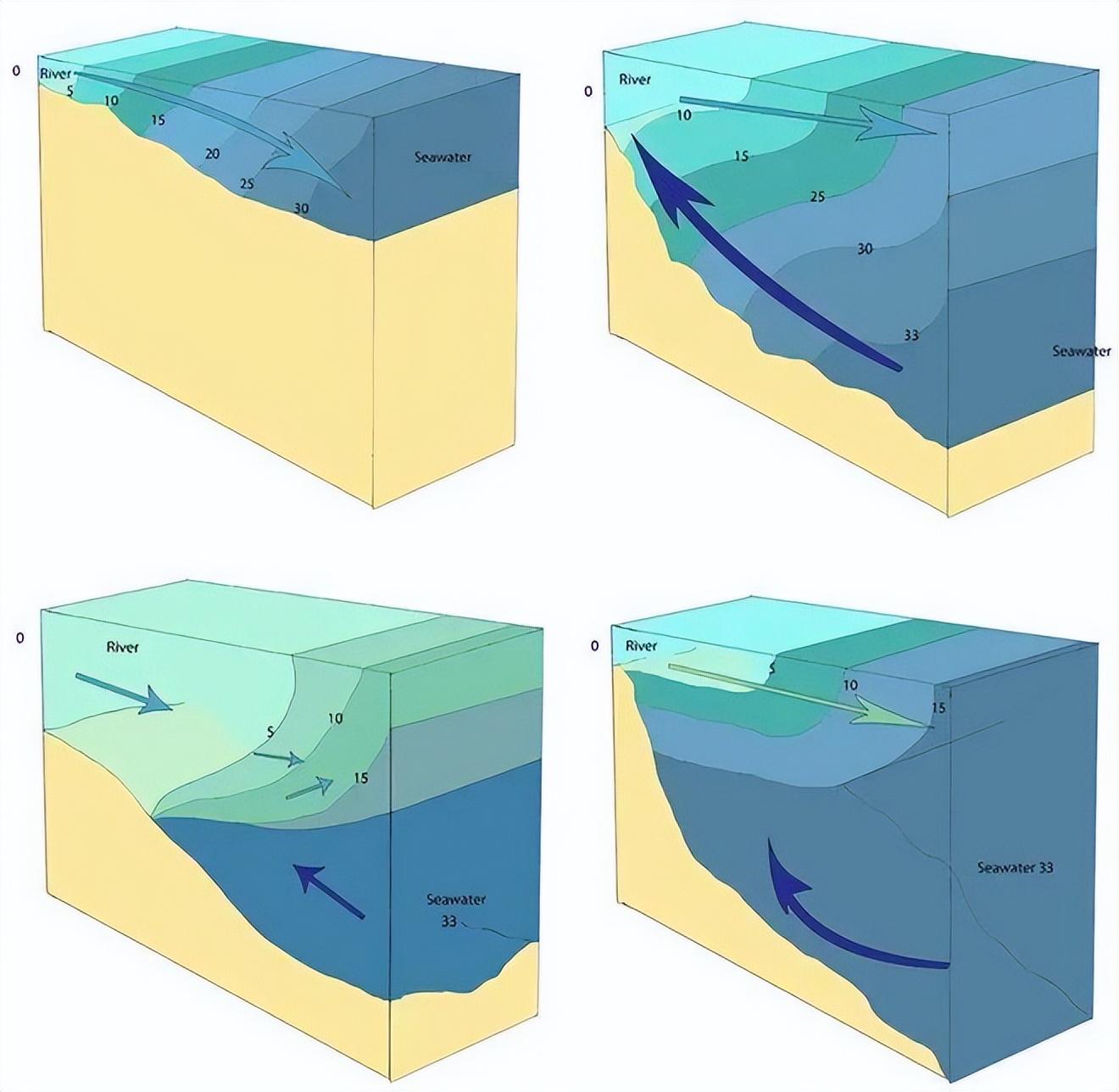
不同河口類型(坡度差異)的鹽梯度變化。來源:libretexts.org
因此其形狀往往呈現舌狀延伸,因此又被稱爲“淡水舌”或“低鹽水舌”。
淡水舌的表層面積與河流日流量正相關,流量超過 200m³/s,淡水舌的面積便能超過 70km²。
而黃河最大流量達到 4700m³/s,平均流量在 2000m³/s左右波動。長江最大流量更是達到 85700m³/s,平均流量約 34000m³/s。黃河具有非常寬闊的淡水舌,長江更是黃河的 10 倍大小。
複雜的水環境,往往吸引不同的海洋/淡水魚類,形成複雜的生態[2]。
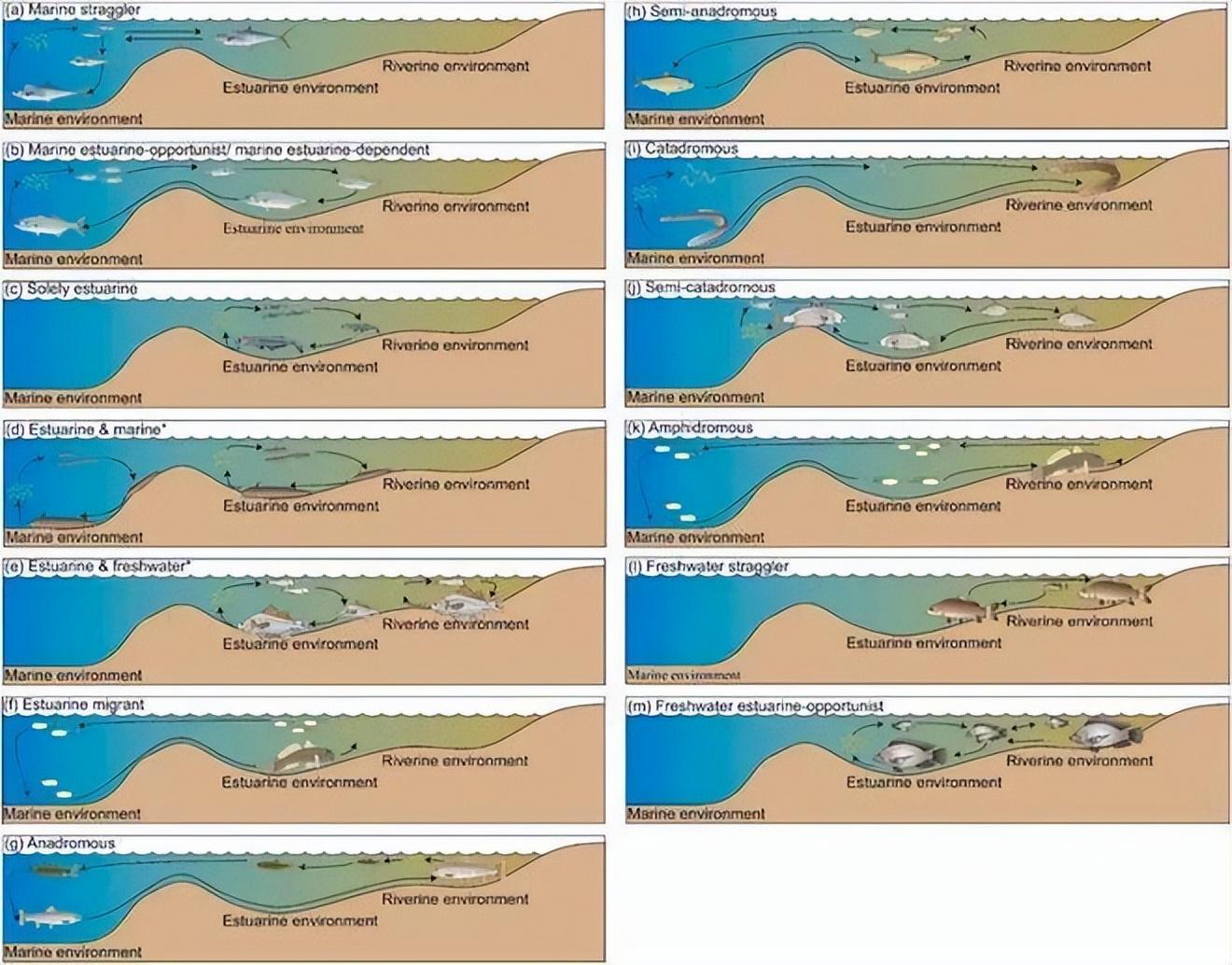
來源:文獻[2]
這裏不僅有誤入的海洋/淡水魚類,偶爾來的機會主義魚類,長期定居的依賴魚類,也有溯河、洄游性魚類。
對於誤闖淡水的典型淡水魚類,例如鯉魚來說,進入大海後,隨着鹽濃度的提升,它們能夠敏銳感知到變化。
因爲它們的味覺感受器不僅分佈在口腔和咽部,還分佈在脣、鰓、須等體表。不僅僅是味覺,甚至它們的嗅覺也有感知鹽濃度的能力,而且具有很高的敏感度。
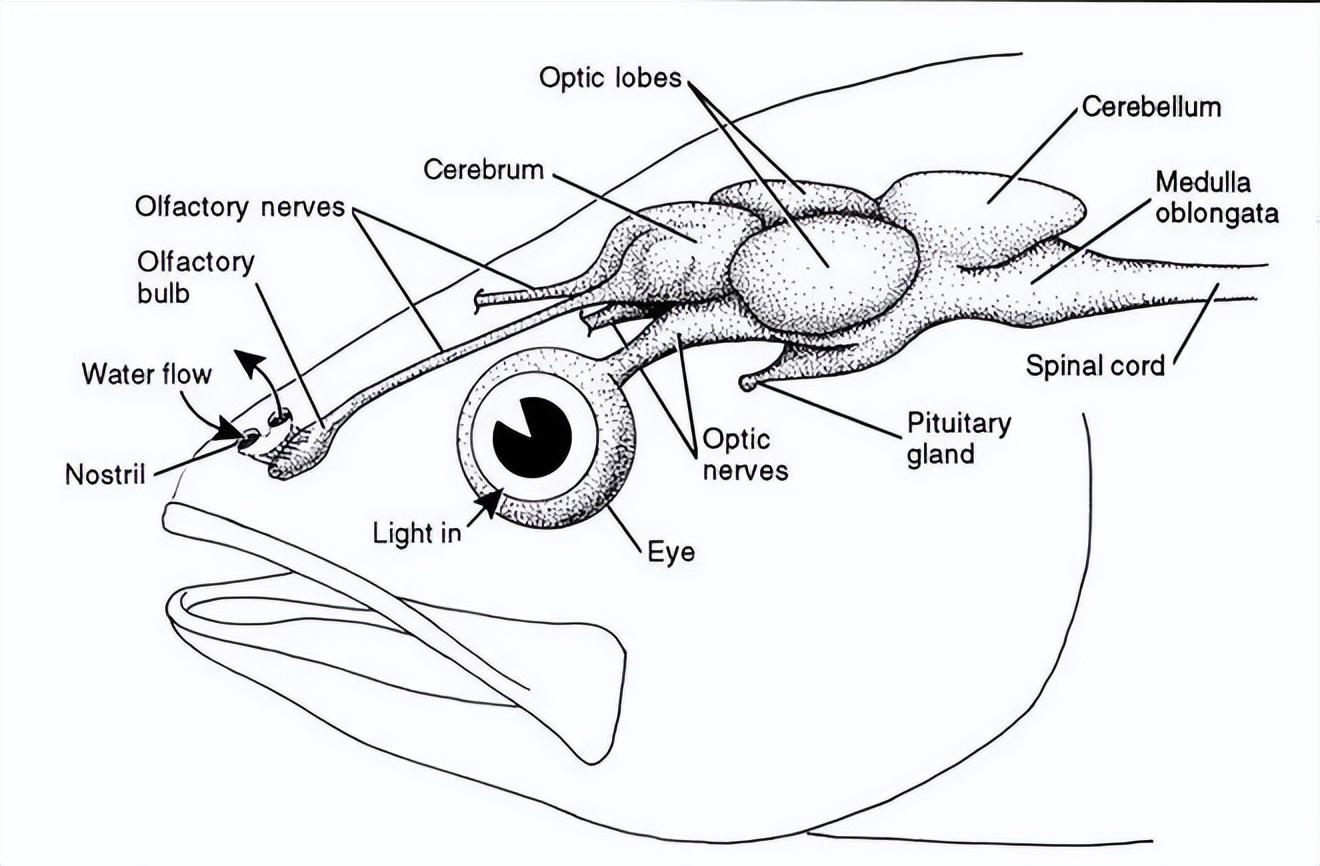
來源:Living Ocean, CRDG, University of Hawaii at Manoa
側線系統雖然沒有直接感知鹽濃度的能力,但可以通過水流密度等性質的改變,間接感知。
當然,還有一個關鍵,那就是魚類直接通過環境中的水和鹽來調節滲透壓[3]。
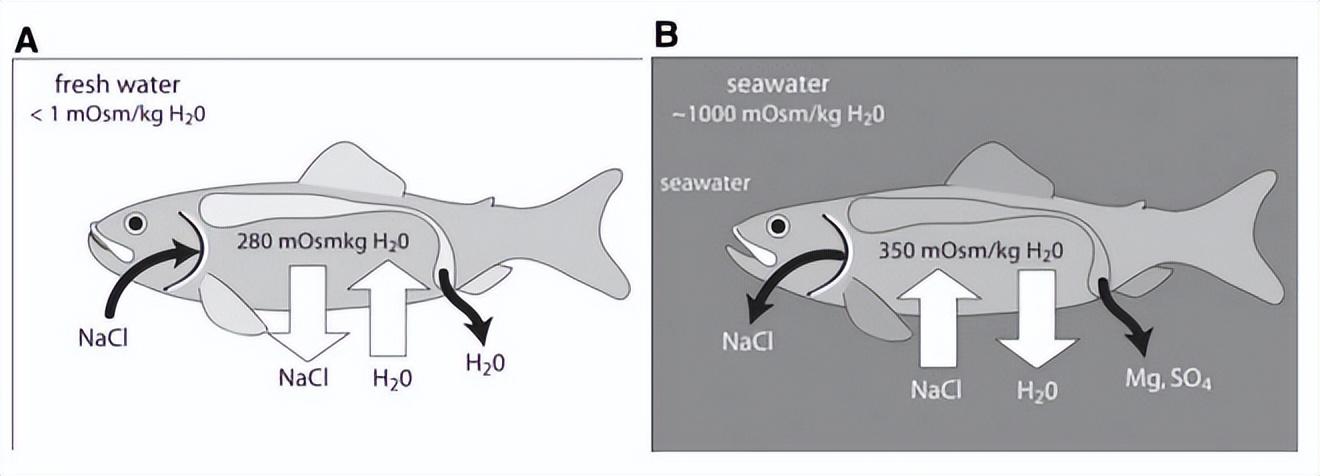
淡水魚(A)和海水魚(B)的滲透壓調節。來源:文獻[3]
隨着環境鹽濃度的改變,魚體會面臨滲透壓失衡。雖然這並不能直接感知鹽分,但不適狀態,與味覺、嗅覺獲得的高鹽濃度信息,可以幫助魚類察覺危險,從而及時回到低鹽水體中去。
一般來說,對於鯽魚、鯉魚這樣的淡水魚,如果能在緩衝區內及時調整狀態,是可以逆流返回長江黃河的。
至於長時間無法回到淡水區的魚類,絕大多數都會死掉。除了因爲滲透壓失衡死掉外,還可能被海洋機會主義掠食者喫掉。
當然,以上分析的是單個魚類個體的情況。
但如果是某種魚經歷漫長的歲月,總是被長期大量衝入入海口,一些存活了下來,那會發生什麼呢?
2
適應環境,掀起一場演化革命
現實中就有活生生的例子,最典型的就是鮭魚。
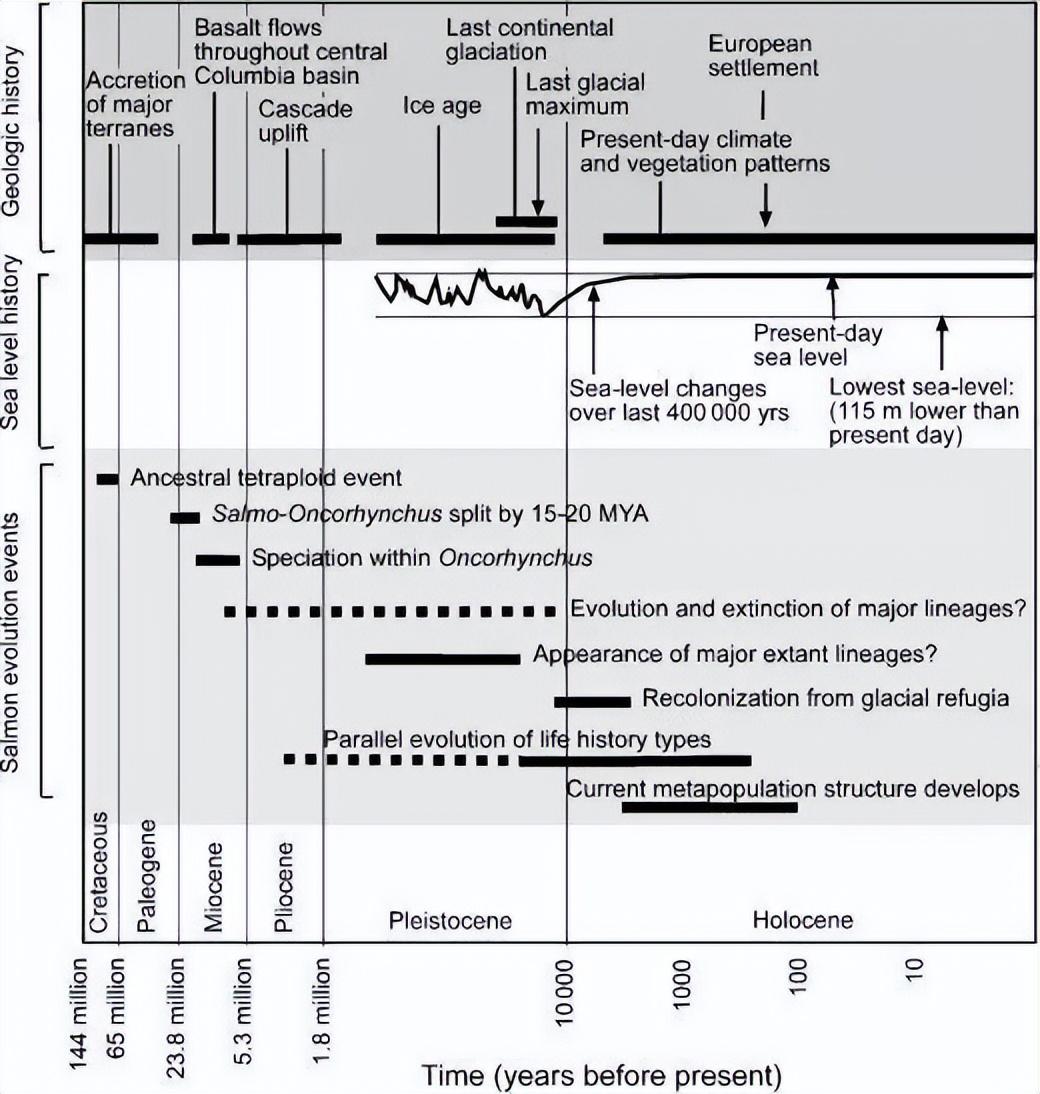
以前認爲鮭魚祖先來自大海,但現在一般認爲來自淡水[4][5],最早 6500 萬~9500 萬年前就開始起源了[6][7][8]。
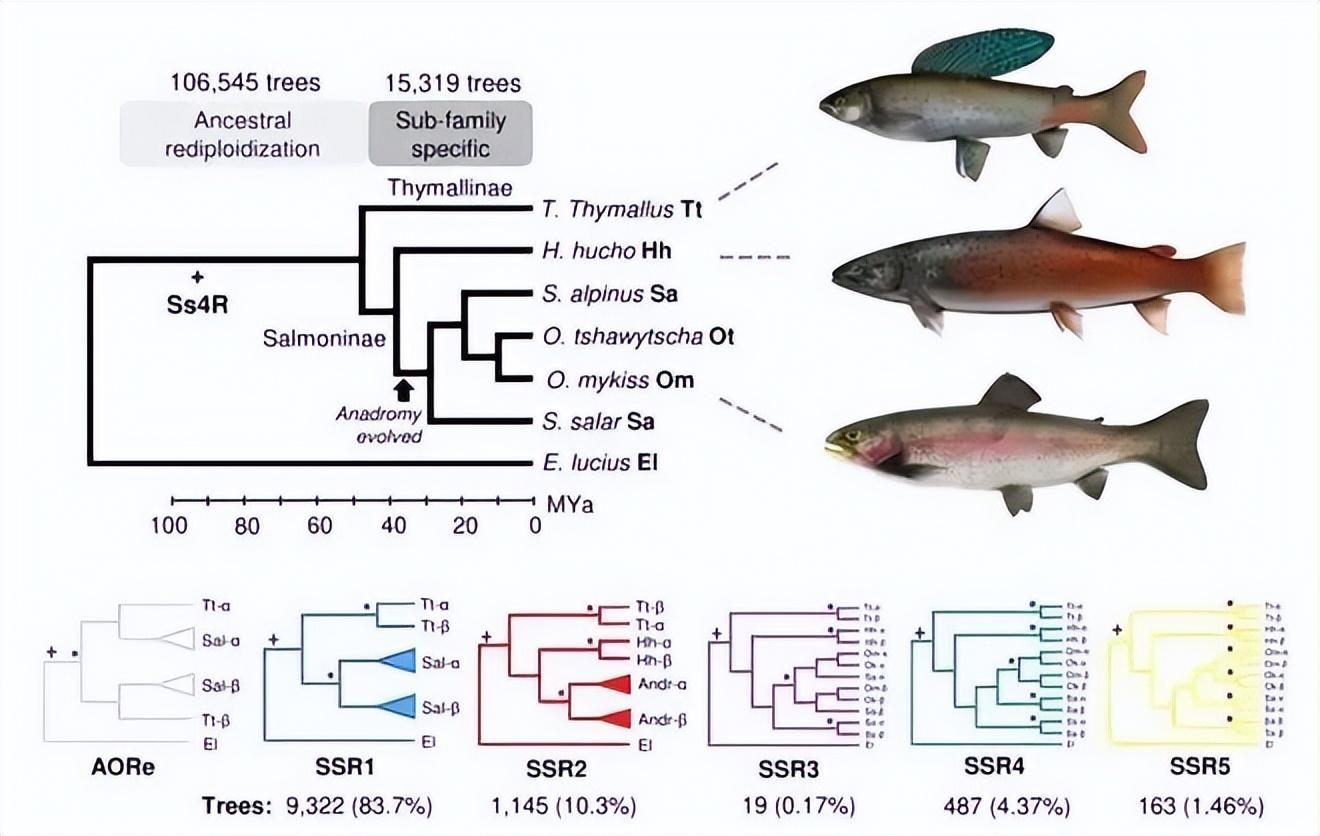
來源:文獻[8]
不斷變化的地理環境,塑造了鮭魚祖先的演化。
3700 萬~5700 萬年前,哥倫比亞河盆地還是廣闊的沿海平原,鮭魚祖先們過得很滋潤。
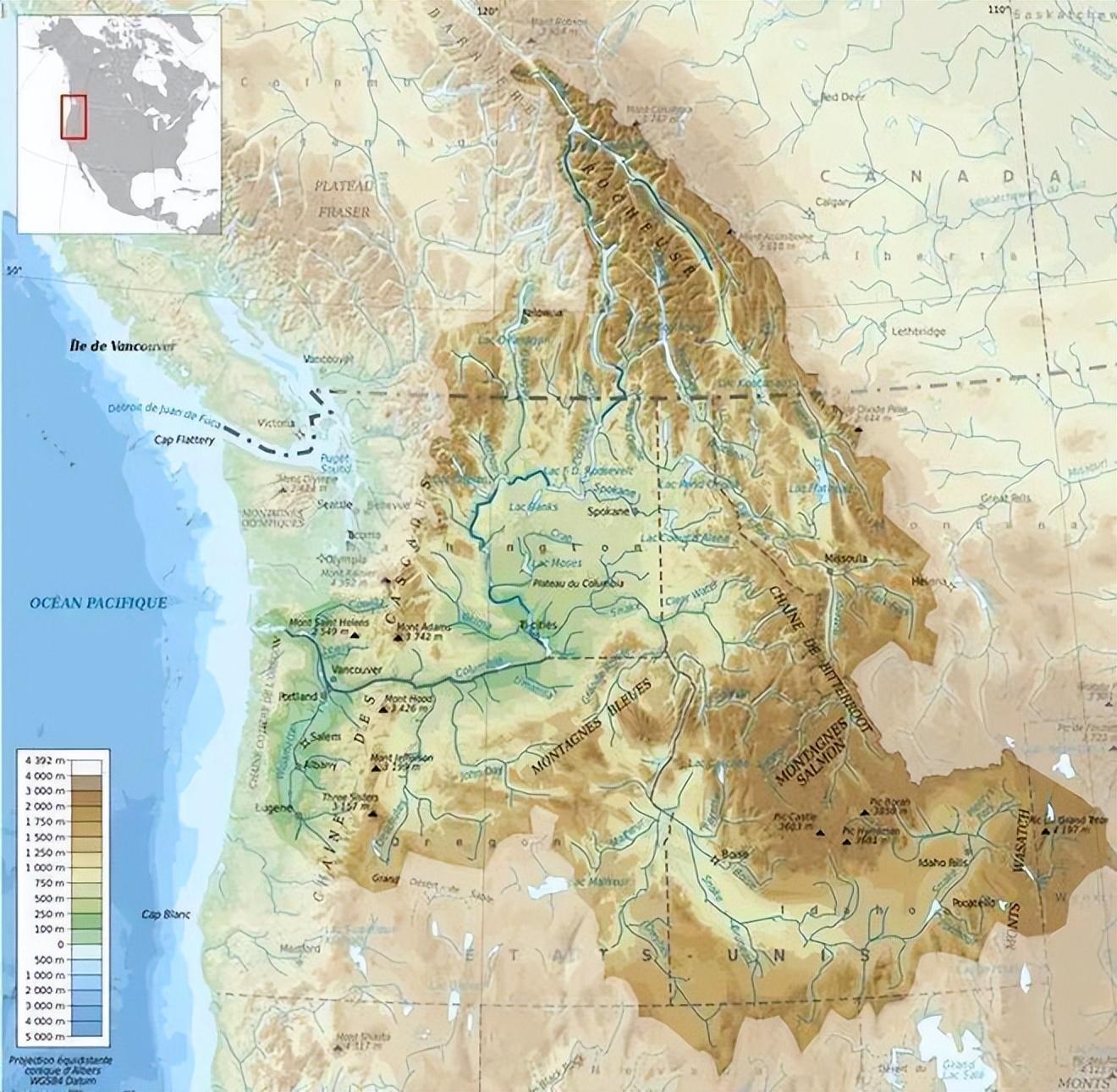
位於北美洲的哥倫比亞河流域。來源:wikimedia
600 萬~1700 萬年前,火山噴發,大規模的玄武岩流過哥倫比亞河盆地中央,最終流入太平洋。不是熔漿就是海水,鮭魚祖先恍如生存於煉獄。
而在 800 萬~1500 萬年間,落基山脈、奧林匹克山脈、以及海岸山脈逐漸抬升,鮭魚祖先們生活的世界,正在改天換地。
來源:文獻[5]
劇烈的環境變化,使得鮭魚祖先適應不同環境的過程中發生了分化。
太平洋鮭魚(Oncorhynchus)的最後祖先,正是從這一時期分化而出。
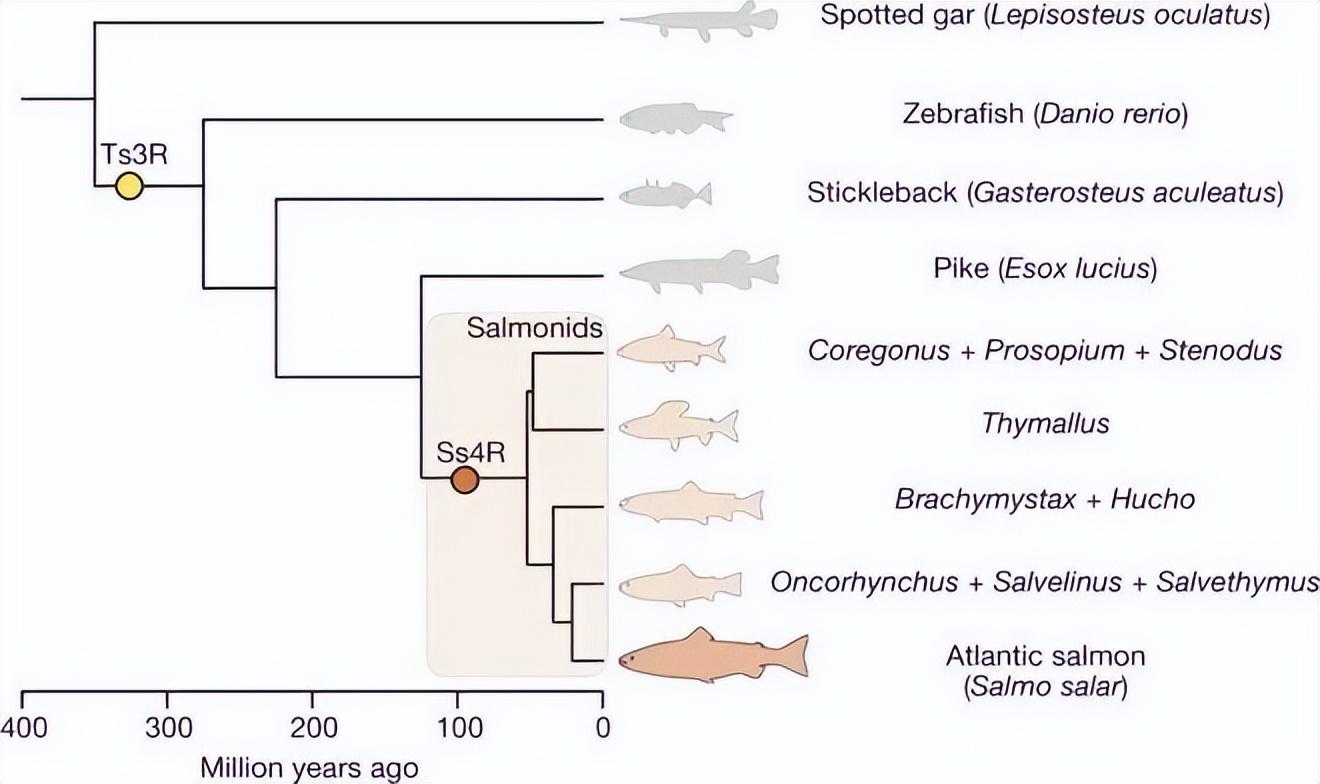
深色區域爲鮭類(Salmonids),鮭科魚類。來源:文獻[7]
最早進入大海的鮭魚科魚類,也可能正好來自 1000 多萬年前[9]。它們有可能在過去幾千萬年至數百萬年前間,發生過多次獨立的“淡水⇄海洋”反覆演化[10]。

來源:文獻[9]
然而,258 萬年前,地球開啓了第四紀冰期。
第四紀冰期並不是一成不變的冷,而是存在大量持續 1~4 萬年的間冰期。
反覆的溫冷交替,使得鮭魚祖先迎來“鬼畜”魚生。
在第四紀冰期期間,多達 19~20 次的反覆冰川作用、溫冷交替、旱洪循環,使得鮭魚在不斷淘汰中演化。
不少早期淡水鮭魚科成員都滅絕了,而太平洋鮭魚經歷不斷被淘汰,適應了冰期的反覆作用。
2 萬年前,在美洲大陸上,一羣太平洋鮭魚熬過了第四紀冰期,開啓了輻射發展的巔峯,終於迎來了生存的曙光。
但它們並不知道,隨着冰期結束,它們面對的是更加“鬼畜”的魚生。
隨着全球變暖,冰壩週期性破裂,發生週期性的洪水。
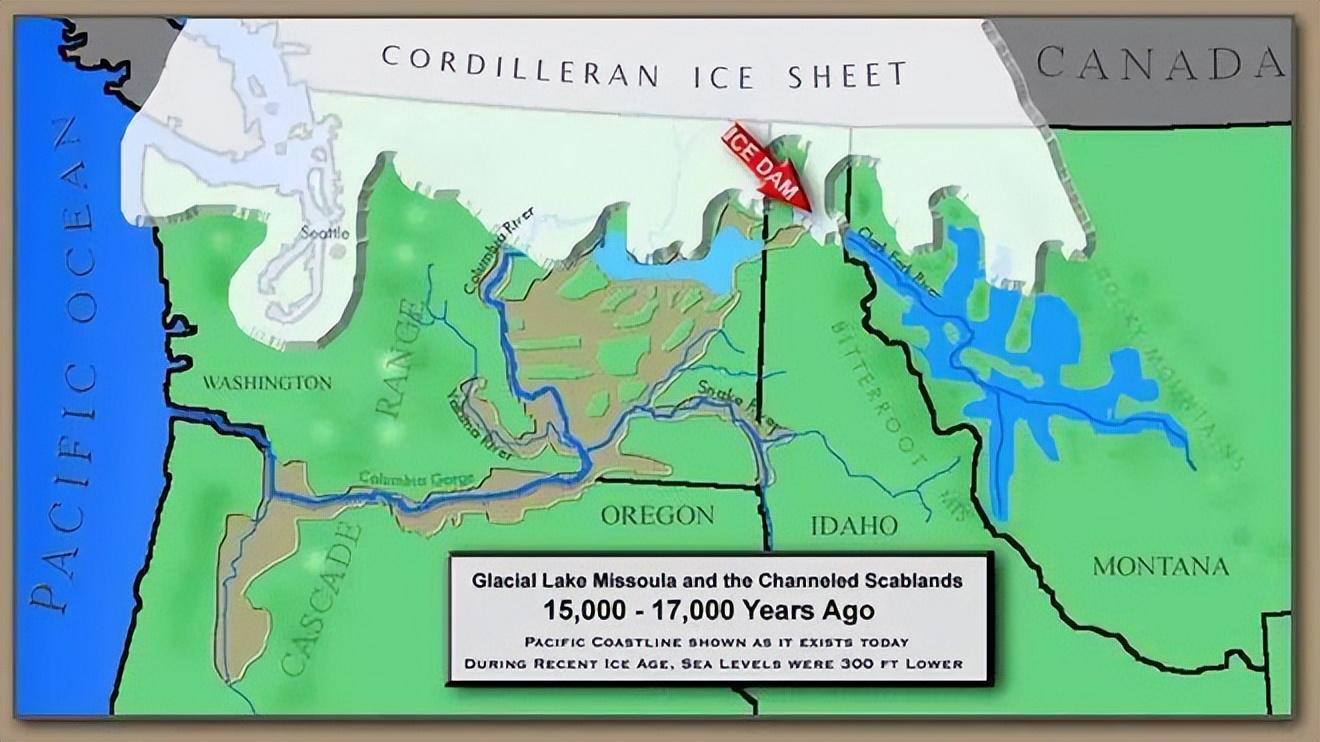
北美大陸 1 萬多年前開啓的冰洪時代。來源:hugefloods.com
冰川和海水的交替作用,一直持續了數千年,其中米蘇拉湖的冰洪就沖刷了哥倫比亞高原上百次。
米蘇拉湖冰洪覆蓋廣闊,流量相當於今天 60 條亞馬孫河,嵌入在冰層中的汽車大小的巨石,被冰洪攜帶,足足漂了 800 公里。
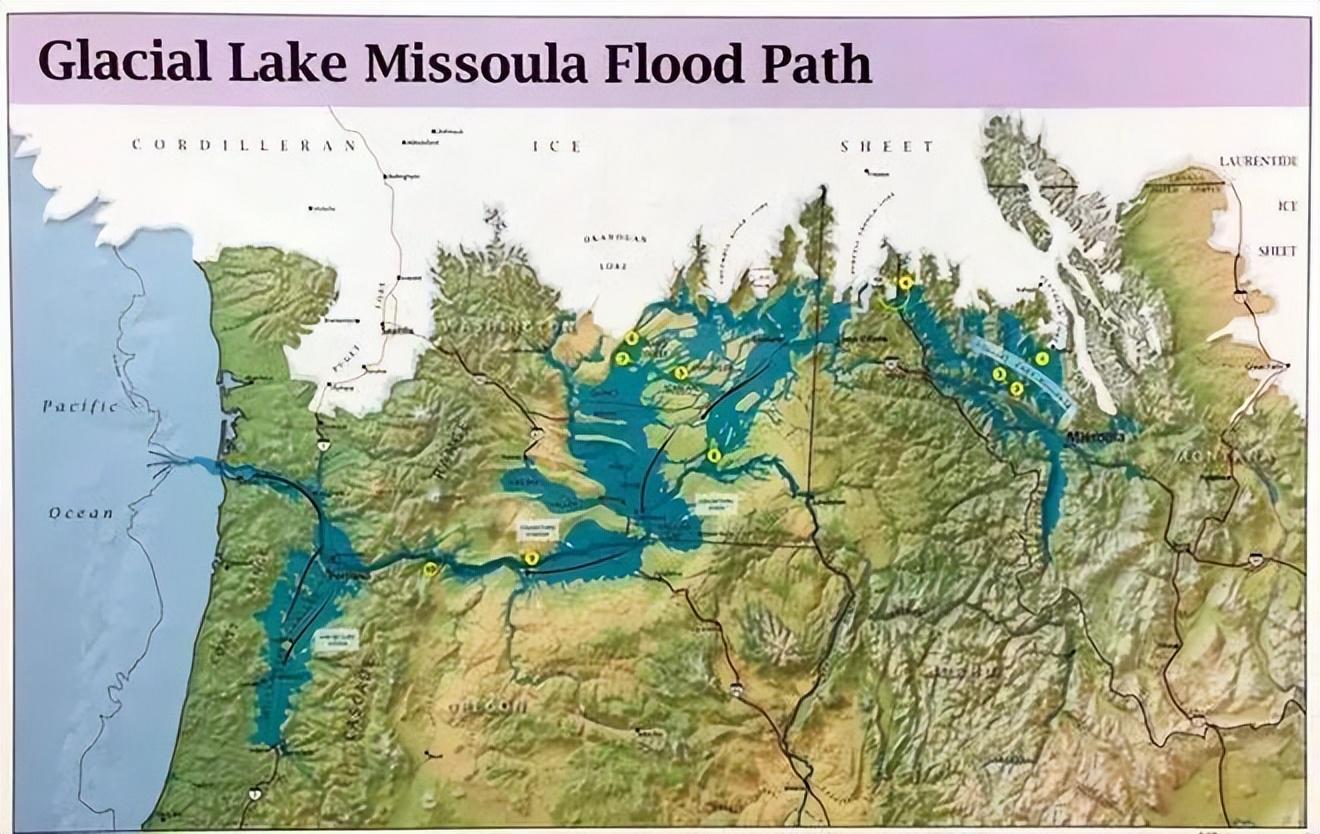
米蘇拉湖冰洪覆蓋面積(左)。來源:Montana Natural History Center
活下來的鮭魚,選擇冰川還是海水?很明顯,從演化的角度來說,在海水中活下來的代價,比在冰川中的代價低很多。
我們可以想象,洄游鮭魚演化的過程。
從第四紀冰期開始到今天的漫長過程,這些鮭魚祖先遭遇冰冷洪水,先死掉了絕大多數。隨後部分進入海水,剩下的部分又死掉了絕大多數。最終,一部分體液滲透壓稍高的個體,扛過一段時間後,再次回到了淡水,從而得以繁衍生息。(由於祖先可能已經發生過“淡水⇄海洋”適應過程,它們再次適應海洋環境也更加的容易。)
當它們發展到足夠規模以後,再次遭遇冰冷的洪水。
經過不斷反覆的淘汰選擇,經過 N 次迭代後,最終活下來的鮭魚,體液滲透壓越來越高,介於淡水魚和海魚之間,甚至還發展出了用鰓排鹽的能力。
它們不僅在海水中存活的時間變長了,而且由於生存淘汰率高,它們也演化出了非常強的繁殖能力。通過足夠高的繁殖數量,來平衡淘汰率高的問題。
同時,要保證足夠多的後代存活下來,也意味着需要洄游到上游,選擇食物充足、天敵少的區域。甚至,爲了後代的生存,不惜犧牲掉自己。
就這樣,經過祖先上千萬年的漫長自然選擇,頂住後來第四紀冰期的“鬼畜”式壓力,又經歷上萬年乃至數千年的激烈冰洪淘汰,最終促使了太平洋鮭魚等鮭魚洄游習性的誕生。
一般認爲,太平洋鮭魚 2 萬年前輻射發展到了巔峯,而洄游習性則至少 11500 年前就存在了[11]。
或許,正是因爲過於殘酷的演化過程,才讓鮭魚的繁殖,達到了自然界少有的高度內卷。
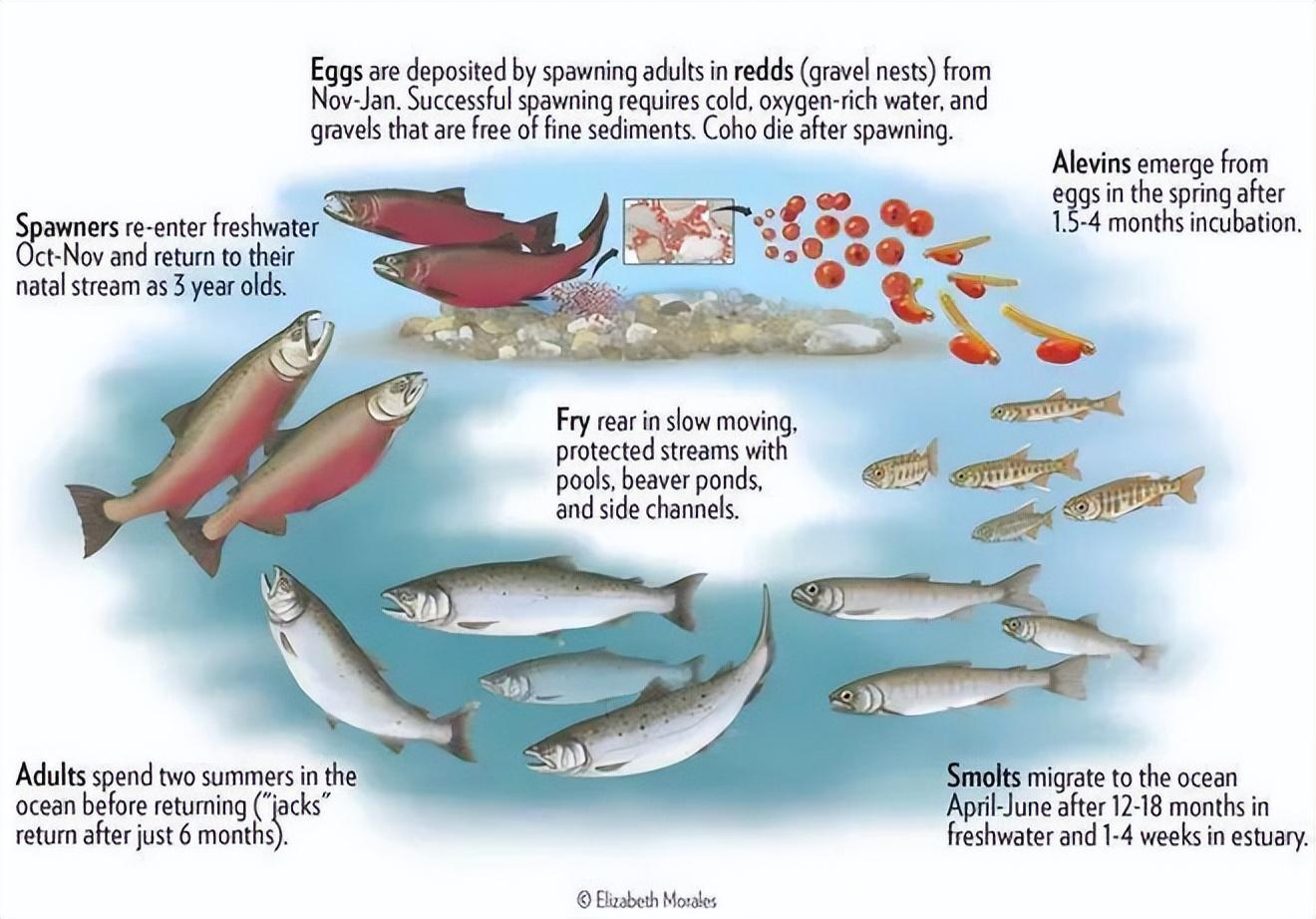
鮭魚生命週期。來源:Elizabeth Morales
鮭魚父母橫穿 800 公里,以死亡爲代價,終生產卵一次。
平均 4000 顆,大約只有 1/5 孵化,最終 200 條入大海,10 條成功洄游,但最終只有 2 條到達出生地,正好一代代的不斷循環。
鮭魚會在河流中生存 1~5 年纔會進入大海,它們成長的保證,正是可以棲息的窪地、河牀、河灣、水草、湖泊……
像鮭魚這樣進入大海,還能活下來的魚類,其實是比較罕見的,需要複雜而特殊的演化壓力。
絕大多數被衝入海水的,除了部分再次回到淡水的,其它的自然是死了,再次進入自然大循環。
你眼裏波瀾壯闊的自然景象,在它們的世界,很有可能是一場殘酷的生存選擇。
參考文獻
[1]Lofi, Johanna, et al. "Fresh-water and salt-water distribution in passive margin sediments: Insights from Integrated Ocean Drilling Program Expedition 313 on the New Jersey Margin." Geosphere 9.4 (2013): 1009-1024.
[2]Whitfield, Alan K., et al. "Importance of marine-estuarine-riverine connectivity to larvae and early juveniles of estuary-associated fish taxa." Environmental biology of fishes 106.10 (2023): 1983-2009.
[3]Beyenbach, Klaus W. "Kidneys sans glomeruli." American Journal of Physiology-Renal Physiology 286.5 (2004): F811-F827.
[4]McDowall R M. The origin of the salmonid fishes: marine, freshwater... or neither?[J]. Reviews in Fish Biology and Fisheries, 2002, 11: 171-179.
[5]Waples, Robin S., George R. Pess, and Tim Beechie. "Evolutionary history of Pacific salmon in dynamic environments." Evolutionary Applications 1.2 (2008): 189-206.
[6]Ramsden S D, Brinkmann H, Hawryshyn C W, et al. Mitogenomics and the sister of Salmonidae[J]. Trends in Ecology & Evolution, 2003, 18(12): 607-610.
[7]Lien, Sigbjørn, et al. "The Atlantic salmon genome provides insights into rediploidization." Nature 533.7602 (2016): 200-205.
[8]Gundappa, Manu Kumar, et al. "Genome-wide reconstruction of rediploidization following autopolyploidization across one hundred million years of salmonid evolution." Molecular Biology and Evolution 39.1 (2022): msab310.
[9]Wood, Chris C., et al. "Recurrent evolution of life history ecotypes in sockeye salmon: implications for conservation and future evolution." Evolutionary Applications 1.2 (2008): 207-221.
[10]Alexandrou, Markos A., et al. "Genome duplication and multiple evolutionary origins of complex migratory behavior in Salmonidae." Molecular phylogenetics and evolution 69.3 (2013): 514-523.
[11]Halffman, Carrin M., et al. "Early human use of anadromous salmon in North America at 11,500 y ago." Proceedings of the National Academy of Sciences 112.40 (2015): 12344-12348.
策劃製作
作者丨瞻雲 科普博主 科普作家 中國科普作家協會會員
審覈丨劉雅丹 中國水產學會原祕書長助理、研究員,全國首席科學傳播專家
策劃丨鍾豔平
責編丨鍾豔平
審校丨徐來、張林林