蟬:從一億年前的“啞巴”到現代的噪音製造者,我經歷了什麼?
蟬鳴作爲夏日的標誌之一,刺耳的聲音可能是所有蟲鳴中最令人煩心的一種了。但沒有蟬鳴的夏季又彷彿是不完整的——這種感覺可能不止中國人有,全世界很多地方的人都會有。蟬科在全球分佈極爲廣泛,其下有超過3000個物種,因此可能全球大部分生活在熱帶-溫帶地區的人都需要適應年復一年的蟬鳴。
雖然對我們來說,蟬鳴是刺耳的,但在昆蟲學家耳中卻非常多樣,猶如“交響樂”一般。“嘒嘒、知了、嗡嗡、鳴鳴、嘶嘶……”,分辨蟬鳴是昆蟲學家必修的課程,因爲他們需要依靠蟬鳴來對蟬科的成員進行分類鑑定。當然,除了用來鑑定身份,蟬鳴對蟬本身也具有重要作用,比如利用蟬鳴識別配偶和防禦捕食者等。
你可能不知道的是,雖然蟬是目前最“大聲”的昆蟲(最高可以發出近120分貝的聲音),但根據化石研究,它們的祖先卻在沉默中度過了上億年時光,以至於跟它們一起生活過的恐龍可能從未聽到過它們的聲音。

蟬總科物種照片。第二行第一張爲螽蟬屬於螽蟬科,其餘都屬於蟬科。
(圖片來源:Marshall et al., 2018)
一億年前,幼蟬的平凡地下生活
時間倒流至約一億年前的白堊紀緬甸熱帶雨林。繁茂的森林沉浸在一片寧靜中,陽光透過樹冠縫隙,將斑駁的光影灑落在覆蓋着苔蘚和蕨類植物的溼潤土壤上。空氣中瀰漫着泥土和植被的清新氣息,時而傳來清脆的鳥鳴聲,時而聽到小型恐龍低沉而急促的呼吸聲伴隨着穿行時奔跑的步伐。
在平靜地表的掩藏下,豐富的生命活動正在土壤世界中悄悄展開。昏昏暗暗的地下世界裏,大量蟬幼蟲們正悠然地沉浸在土壤的柔軟之中。這些約1億年前白堊紀的蟬幼蟲呈現出與現代蟬幼蟲相似的形態特徵,最引人矚目的便是它們鐮刀狀的脛節與擴張膨大的股節相契合形成抓握結構的特化前足。
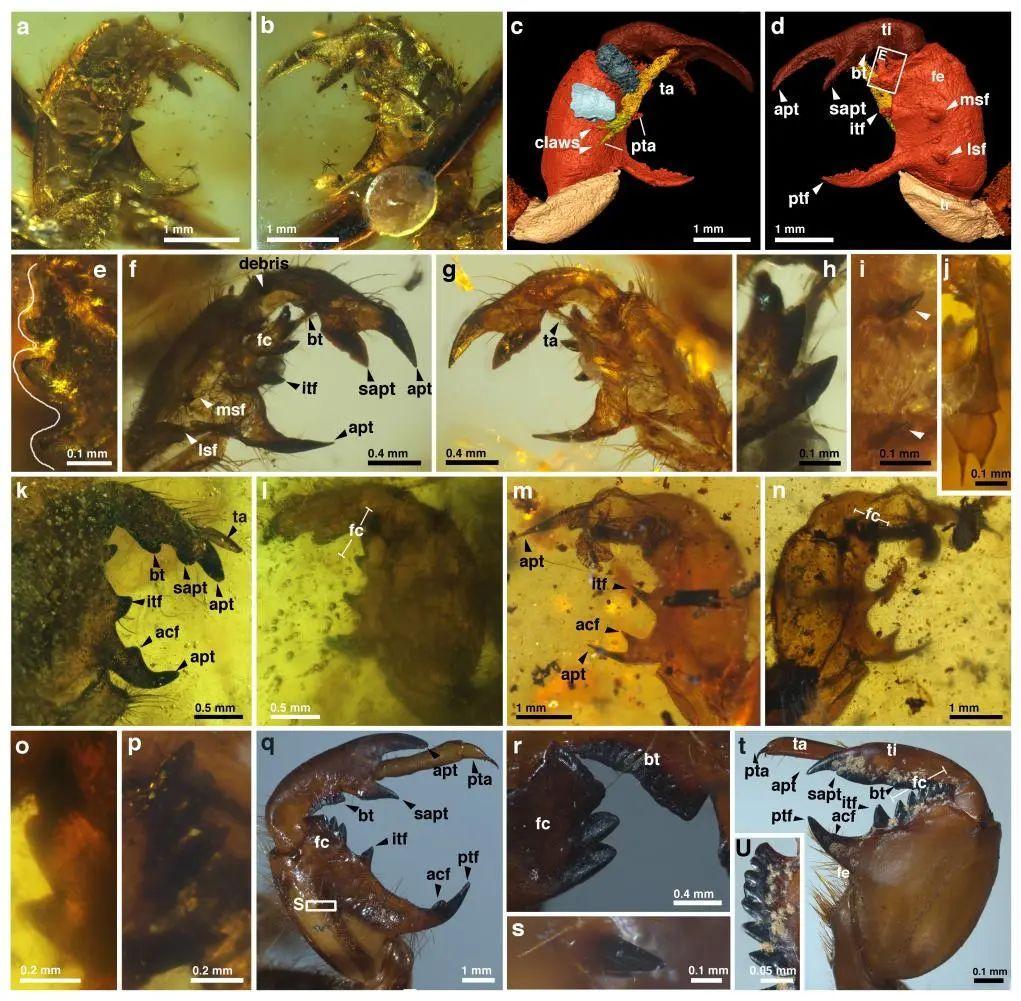
蟬蛻前足化石和現生螽蟬和蟬科物種前足。
(圖片來源:Jiang et al., 2024)
儘管特化的前足讓它們看起來異常兇猛,但蟬幼蟲並無攻擊之心,特化前足僅用來幫助它們擁有更強的土壤挖掘能力,更加適應於地下生活。土壤中隨處可見這些蟬幼蟲的勞作成果:發達的隧道、寬敞的巢穴,還有形成半月狀的沉積物回填體。

左圖上方爲現代蟬幼蟲地下活動模擬實驗,下方爲昆蟲活動遺蹟化石對比;右圖爲現代蟬生活週期示意圖。(圖片來源:Smith et al., 2008)
此時一隻蟬幼蟲的特化的前足正微微向內傾斜,在巢穴中交替地挖掘沉積物並向前移動。一塊沉積物被鏟向它的頭部,並壓實成球狀團塊夾在頭部和前腿之間,一個180度前滾翻,沉積物被帶到了身後,而後它再用前足將沉積物推向巢穴後方。
周圍大部分蟬幼蟲此刻正在它們的巢穴裏休息或者進食。它們的巢穴通常建造在植物的根部附近,蟬幼蟲用自己口器中的口針刺入植物根的木質部,脣基配合地吮吸其中的汁液用來果腹,體內的共生菌也在賣力幫它們合成所需的營養成分。這些蟬幼蟲還將自己的排泄液體變廢爲寶,用作“自制膠水”黏合土壤,打造出堅固的地下巢穴。

左圖蟬幼蟲巢穴遺蹟化石;右圖現代蟬巢穴遺蹟。
(圖片來源:Krause et al., 2008)
就這樣,億年前的雨林土壤下靜謐而祥和,生命以其輕盈的步伐,悄悄譜寫着自然的故事。對蟬幼蟲而言,時間在地下慢慢流淌,不知不覺可能就在地下生活了數年。現代的蟬幼蟲最長可在地下生活長達17年。

約1億年前白堊紀中期緬甸琥珀中末齡蟬幼蟲化石。左側化石照片,右側顯微CT數據三維重建。
(圖片來源:Jiang et al., 2024)

約1億年前白堊紀中期緬甸琥珀中蟬蛻化石。
(圖片來源:Jiang et al., 2024)
幼蟲到成蟲的蛻變
隨着夏日的來臨,地面溫度的累積達到某個觸發點,從天色漸暗的傍晚直至月光柔和的午夜,蟬幼蟲一個個小心翼翼地從地面破土而出,彷彿是自然界精心策劃的一場盛大儀式。蟬幼蟲到地面後,便開始尋找適宜之地,準備進行一生中重要的蛻變,以邁入生命的新階段。它們的背部外殼開始緩緩裂開,露出新生的、柔軟的成蟲身體。而新生的成蟲則需要用盡全力,慢慢從舊殼中掙脫出來。
經過一段時間的等待,這些剛完成蛻變的白堊紀成蟲終於展現出它們的真實面貌。就在這一瞬間,時間彷彿被落下的一滴琥珀定格,琥珀巧妙地捕捉並封存了億年前的生命場景。緊接着,鏡頭彷彿又被時間的手輕輕撥動,畫面從古老的熱帶雨林穿梭回現代,最終聚焦在實驗室顯微鏡的鏡頭下。

約1億年前的白堊紀中期緬甸琥珀中成蟲蟬化石。
(圖片來源:Jiang et al., 2024)
在鏡頭下觀察,會發現這些成蟲與我們熟悉的會發聲的蟬科成員並不完全一樣,似乎兼顧了蟬科和其姊妹類羣螽蟬科特點。與龐大的蟬科家族不同,螽蟬科僅分佈在澳大利亞,包含1屬2種,被認爲是蟬科僅存的近親。螽蟬科和蟬科共同組成一個家族,稱爲蟬總科(超科級別)。

左圖螽蟬,屬於螽蟬科。(圖片來源:https://www.inaturalist.org) 右圖Greengrocer蟬,屬於蟬科。(圖片來源:Australian Museum)。
螽蟬科的顯著特點是擁有一個巨大的前胸背板,前胸背板遮蓋了絕大部分中胸背板,只露出末端三角狀小盾片。而蟬科成員大多展示出明顯的中胸背板,其末端形成了特有的十字隆起結構。
爲了探究琥珀中的這些化石蟬與現代蟬科及螽蟬科之間的關係,中國科學院南京地質古生物研究所的古生物學家們採用了顯微CT技術來對標本進行解析。這一技術使研究人員能夠無損地對白堊紀時期以及現代蟬的成蟲和幼蟲身體結構進行詳細檢查。

南京地質古生物研究所顯微CT實驗室。
(圖片來源:吳素萍攝)
實驗過程是這樣的:首先研究人員將標本放置於顯微CT儀器內,調試好參數後,樣品便開始進行360°的旋轉掃描,以獲得一系列X光片。接着,利用三維視圖軟件對這些X光片進行重構,最終生成了蟬的3D圖像,從而收集了標本一系列豐富的形態特徵數據。
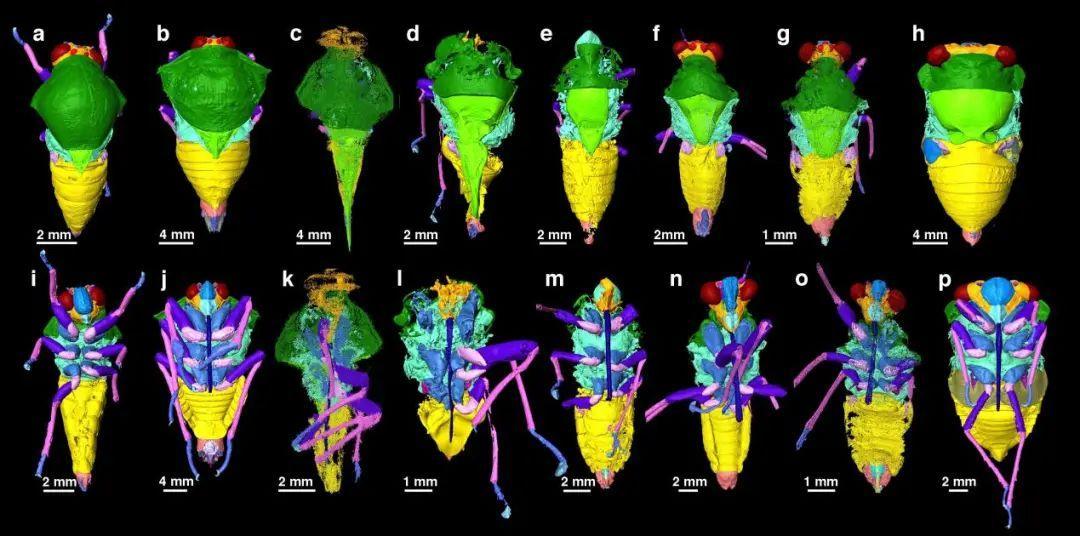
琥珀中成蟲蟬化石和現生類羣顯微CT數據三維重建。
(圖片來源:Jiang et al., 2024)
此外,通過翻閱化石家譜記錄,研究人員還獲得了1.65億年前侏羅紀時期蟬親戚們的形態特徵,並將這些數據一同納入系統發育和形態差異分析,以瞭解化石蟬與現代蟬科及螽蟬科之間的關係。

1.65億年前侏羅紀時期蟬總科化石。
(圖片來源:南京地質古生物研究所)
分析結果表明包含現代蟬科和螽蟬科的兩個祖先譜系至少在侏羅紀中期就已經分道揚鑣了。這些侏羅紀和白堊紀的蟬化石都屬於蟬總科家族,其中一些過去被認爲和螽蟬親緣關係較近的化石,現在被認爲與蟬科關係更近,這大大增加了我們對蟬科起源與演化的認識。隨着蟬總科內部的親緣關係的梳理和明確,一個引人入勝的問題浮現出來:這些白堊紀時期的蟬祖先親戚們,是否也能像今天的蟬科一樣,發出獨特而響亮的叫聲呢?
一億年前,蟬還不會發出響亮的鳴叫
現代蟬發出鳴聲通常與腹部鼓膜結構有關。經過顯微CT檢測和化石三維重建,古生物學家發現,這些琥珀中屬於蟬總科家族成員的化石蟬保存了鼓膜結構,且雌性和雄性化石中均有保存鼓膜結構。現存螽蟬科成員和絕大多數蟬科成員也都具有鼓膜結構。因此,結合現代研究結果,這些琥珀中的化石證據證明了具有鼓膜結構是蟬總科家族的一個早期原始特徵,並被代代遺傳了下來。那麼,這些化石蟬保存有鼓膜結構能否表明它們也具有發出響亮鳴聲的能力呢?
現存蟬科中的鳴蟬主要是雄性來發出高亢的聲音。雄蟬利用腹部發達的鼓膜肌肉運動通過表皮內突來牽引鼓膜,致使膜上面的肋來回彎曲產生聲音。它們的腹部具有共振腔,共振腔像一個天然的音響,進一步放大這些聲音,在空氣中傳播以實現交流。當然鳴蟬還發育了聽覺系統,包括聽膜、聽膜脊和聽囊等結構,來配合接收聲音。
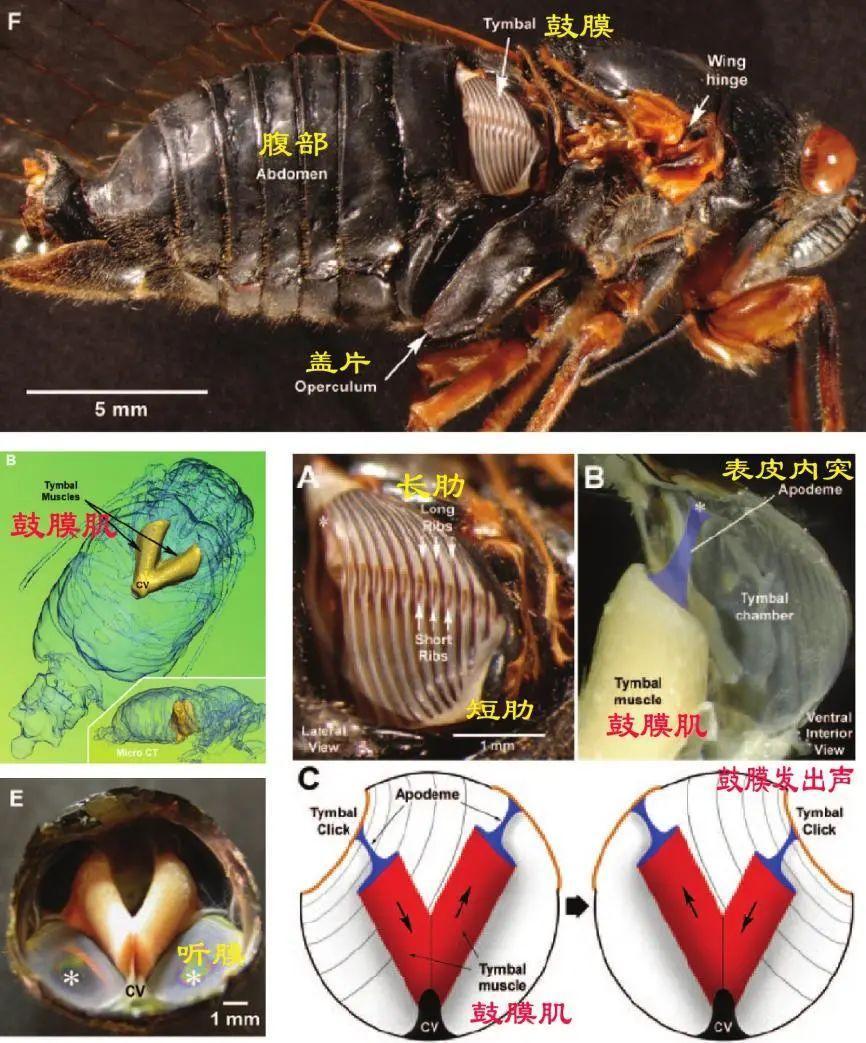
(圖片來源:Nahirney et al.,2006)
與鳴蟬不同,螽蟬很安靜,不會發出響亮的鳴聲。它們也類似鳴蟬,通過腹部運動產生振動信號,但產生的振動信號則是通過樹幹樹枝等固體基底進行傳遞,使用腿部感受器接收信號,從而實現交流。儘管螽蟬也有鼓膜和鼓膜肌,但並不發達,並且腹部缺乏共振腔。
可見,蟬發出響亮的鳴叫聲不僅依賴鼓膜,還要依靠腹部整套發聲裝置的配合。僅僅發現鼓膜,並不能說明化石蟬是否能發出響亮的鳴叫。對琥珀中的化石蟬進一步身體檢查發現,大多數化石蟬並未發現其他與發聲和聽覺相關的結構,或者明顯發育且類似於現代鳴蟬的結構特徵。
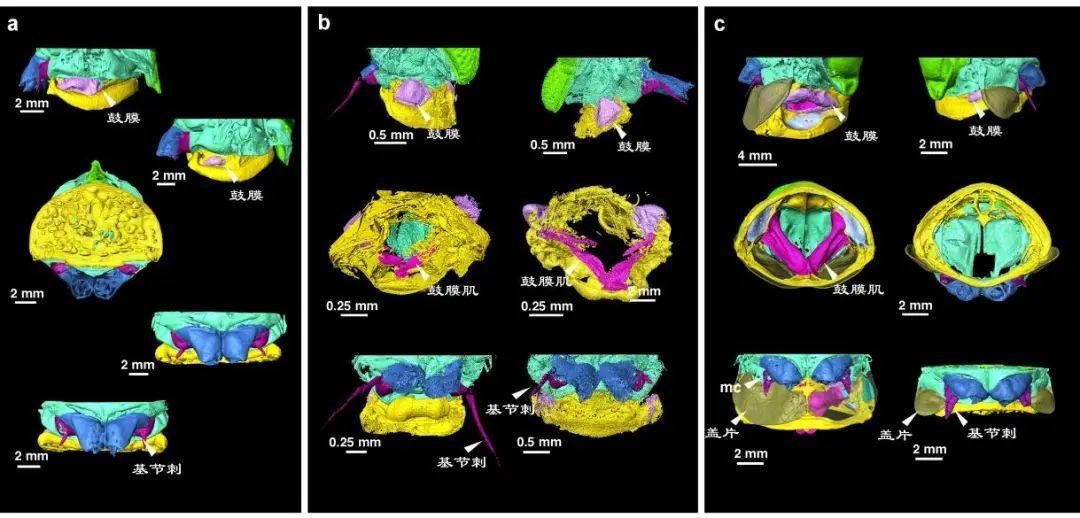
顯微CT數據三維重建,現代蟬與化石蟬腹部結構比較。左邊雄性,右邊雌性。a. 螽蟬(螽蟬科,現代標本);b. 夏氏原始蟬(化石); c. 蟪蛄 (蟬科,現代標本)。(圖片來源:修改自 Jiang et al., 2024)
由此古生物學家得出結論,這些生活在約1億年前的白堊紀中期的蟬總科成員可能沒有複雜的發聲和聽覺裝置,無法產生和接收類似現代鳴蟬的高分貝歌聲。它們的溝通方式可能與今日的螽蟬更加相似,身體產生振動信號,通過枝葉傳遞,悄悄地交換心聲。

Pranwanna xiai 夏氏原始蟬。
(圖片來源:南京地質古生物研究所)
此外,還有一隻小化石蟬“夏氏原始蟬”,大約1釐米長,不僅保存了鼓膜結構,還保存了較爲發達的鼓膜肌肉和明顯的腹部空腔。這一結構,配合其氣管、飛行肌等身體內部結構被完整地以化石形式保存下來,似乎表明這隻化石蟬腹部空腔本身就存在,很有可能具備了共振腔。
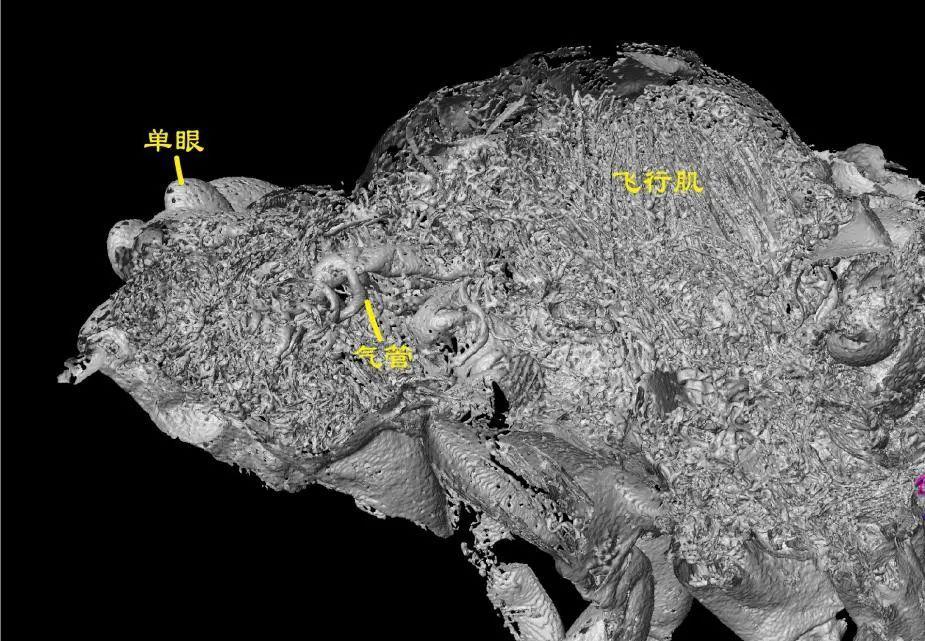
夏氏原始蟬化石中器官、飛行肌結構。
(圖片來源:Jiang et al., 2024)
夏氏原始蟬也代表了目前在白堊紀中期緬甸琥珀中發現的與現代蟬科親緣關係最爲接近的蟬總科成員。這似乎向我們透露了另一個有趣的可能性,即在白堊紀中期,一些蟬總科成員可能可以發出比典型的通過固體傳遞的振動信號更明顯的聲音。但即便如此,它的發聲和聽覺系統都無法與現代蟬鳴聲相媲美,始終還是無法演奏出像現代蟬那樣高亢的鳴聲。

中生代森林中蟬的生活生態場景示意圖。
(圖片來源:Jiang et al., 2024;楊定華繪製)
到此,通過研究琥珀化石,科學家們已經將蟬的演化歷程大致梳理了出來。但是當我們從化石研究中抬起頭來,卻又會發現另一個更加現實的問題:這些存在了上億年的“交響樂家”們,在現代由於受到城市化快速擴張、植被過度採伐及氣候變化影響,其全球數量正急劇下降(Yang et al., 2023)。或許,當夏日的蟬鳴再次響起時,那不僅僅是大自然的交響曲,也是一份溫柔的邀請——珍惜當下,攜手保護,確保蟬的歌聲在未來繼續悠揚,而非僅存於我們的記憶之中。
參考文獻:
[1]Jiang, H., Szwedo, J., Labandeira, C. C., Chen, J., Moulds, M. S., Mähler, B., Muscente A. D., Zhuo D., Nyunt T. T., Zhang H., Wei C., Rust J. & Wang, B. (2024). Mesozoic evolution of cicadas and their origins of vocalization and root feeding. Nature Communications, 15(1), 376.
[2] Krause, J. M., Bown, T. M., Bellosi, E. S., & Genise, J. F. (2008). Trace fossils of cicadas in the Cenozoic of Central Patagonia, Argentina. Palaeontology, 51(2), 405-418.
[3] Marshall, D. C., Moulds, M., Hill, K. B., Price, B. W., Wade, E. J., Owen, C. L., Goemans, G., Marathe, K., Sarkar, V., Cooley, J. R., Sanborn, A. F., Kunte, K., Villet, M. & Simon, C. (2018). A molecular phylogeny of the cicadas (Hemiptera: Cicadidae) with a review of tribe and subfamily classification. Zootaxa, 4424(1), 1-64.
[4] Nahirney, P. C., Forbes, J. G., Douglas Morris, H., Chock, S. C., & Wang, K. (2006). What the buzz was all about: superfast song muscles rattle the tymbals of male periodical cicadas. The FASEB journal, 20(12), 2017-2026.
[5] Smith, J. J., & Hasiotis, S. T. (2008). Traces and burrowing behaviors of the cicada nymph Cicadetta calliope: Neoichnology and paleoecological significance of extant soil-dwelling insects. Palaios, 23(8), 503-513.
[6] Yang, H., Xiang, X., Wu, X., Thompson, J. R., & Flower, R. J. (2023). Silent summers: The decline of cicadas. Science, 381(6657), 490-490.
作者:姜慧 王博
作者單位:中國科學院南京地質古生物研究所